
Semi-arid ecosystems particularly sensitive to mega-droughts

Drought has affected most regions of the globe in the early 21st-century. In Australia, the recent Millennium Drought from 2001 until 2009 was the worst on record and ended dramatically with one of the largest La Niña associated wet periods in 2010-2012 (Ummenhofer et al., 2011; van Dijk et al., 2013). The early 21st-century drought had significant impact, including significant reduction in agricultural production, reduced water availability, and increased forest die-back and bush fires.
Amplification of the hydrological cycle as a consequence of global warming is predicted to increase climate variability and extreme climatic conditions such as drought or heatwaves are very likely to intensify in coming decades. One of the most urgent and significant questions is whether ecosystems could maintain their health and services, such as carbon accumulation and food production, under intensive climatic extremes impacts.
Writing in Journal of Geophysical Research, we found that drought and wet extremes to strongly affect the vegetation development of ecosystems (Ma et al., 2015). Among different ecosystem types, semi-arid system was most vulnerable to drought, raising concerns on their resilience under future mega-drought. Our research was highlighted the latest issue of Nature.
Monitoring drought stress on vegetation dynamics using ground meteorological measurements and observations from space-borne sensors
We investigated the impacts of the early 21st-century extreme hydroclimatic variations in southeastern Australia on seasonal vegetation development (phenology) and vegetation productivity using remote sensing and meteorological datasets. To measure seasonal vegetation development, we used a spectral vegetation index called enhanced vegetation index, which is calculated using spectral reflectances at visible and infrared bands measured from space-borne sensor and is a composite property of green leaf area, canopy structure, and leaf chlorophyll content. To quantify the extent end intensity of drought, we used a drought index called standardised precipitation and evapotranspiration index, which is calculated using precipitation and temperature data measured from ground stations and reflects the cumulative effect of the imbalance between atmospheric supply and demand of water.
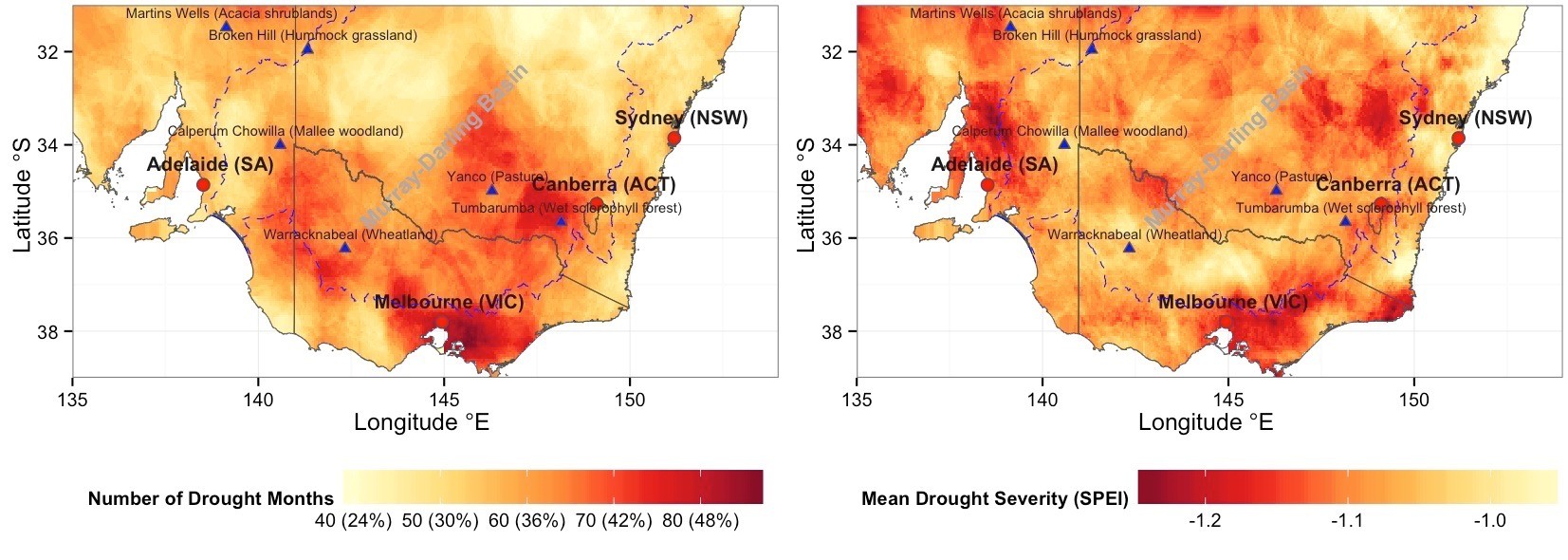
Figure 1 | Drought frequency and drought severity in Southeastern Australia during the 2000-2013 time period. Various “hot-spots”, affected by drought more than other areas during the early 21st-century, were identified, including almost all of southern Victoria and southwestern New South Wales.
Using these datasets, we found that drought and wet extremes have dramatic impacts on vegetation dynamics, with abrupt between year changes in seasonal vegetation development (phenology) and productivity (Figure 2).
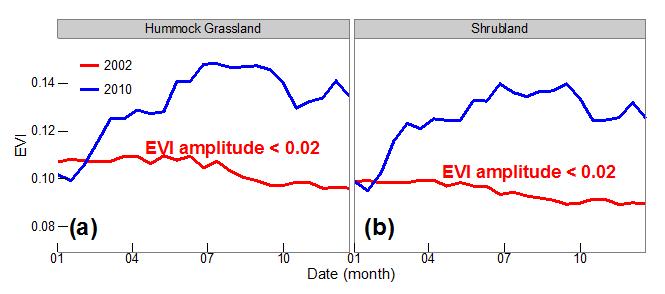
Figure 2 | Comparison of hydroclimatic variation-induced shifts in seasonality for hummock grassland and shrubland, respectively. The solid red and blue lines show the EVI profiles for these two vegetation types in 2002 drought and 2010 wet years, respectively. EVI profiles were averaged using pixels seriously affected by 2002 drought; thus, no phenology was detected during that year. Image modified from Ma et al. (2015).
Abrupt shifts in vegetation phenology and productivity under climatic extremes
Vegetation dynamics and the phenological metrics derived from ground or remote sensing observations for describing these dynamics, e.g., leaf flush or onset of growing season, are key indicators of ecosystem responses to climate variability and change. We found dramatic impacts of drought and wet extremes on phenology and vegetation productivity. In contrast to midlatitude and high-latitude biomes in the Northern Hemisphere that generally have recurrent phenological cycles, driven predominantly by temperature variation, the high interannual variations in vegetation phenology and productivity at Australia’s dryland ecosystems not only highlighted the extreme climatic variability in Australia but also revealed a high phenological and functional plasticity of Australia’s native vegetation.
For instance, vegetation growth of hummock grassland (Spinifex) can be nearly completely dormant during the extreme drought period, yet still maintain capability to be highly productive when favorable periods arrive, reflect the ecological resilience of these vegetation to hydroclimatic extremes (Figure 3). These abilities are essential for Australia’s arid native vegetation to survive and thrive in such a dry and variable climate.

Figure 3 | Australia’s unique native vegetation type Hummock grassland (Spinifex) in dry season (left) and wet season (right). Hummock grassland (Spinifex) covers ~20% of Australia’s dryland. These photos were taken at Ti Tree East OzFLUX tower site north of Alice Springs. Photo Credit: Dr. James Cleverly (UTS: School of Life Sciences).
Semi-arid ecosystems are most vulnerable to the drought, while arid native vegetation exhibit high drought resilience
A crucial question emerges from our research is: which vegetation types and ecosystems are most vulnerable to extreme climatic conditions, and which are most resilient? An additional and unique finding of this study is that ecosystem hydroclimatic sensitivity peaked over semi-arid, instead of more water-limited arid ecosystems. This is not expected as biomes with the largest limitation in water availability were also expected to show the largest sensitivity to hydroclimatic variations. A recent study suggested that the loss of resilience associated with dieback would probably occur first at ecosystems that are most sensitive to precipitation variability (Ponce-Campos et al., 2013). Our finding of hydroclimatic sensitivity peaking in semiarid ecosystems suggests that these semi-arid systems are most vulnerable to climatic extremes and are most likely to experience severe loss of ecosystem resilience with future megadrought events.
The risk of loss of semi-arid carbon sink under a drying trend
Globally, semi-arid ecosystems cover 40% of the global land surface. Australia is the driest inhabited continent in the world, with arid and semi-arid ecosystems together cover nearly 85% of Australia (Figure 4). Recent evidence suggests that global semi-arid ecosystems can dominate inter-annual variability of global carbon cycle (Poulter et al., 2014; Ahlström et al., 2014). For example, an exceptionally large global land carbon sink anomaly was recorded in 2011, of which more than half was attributed to Australia’s semi-arid ecosystems.
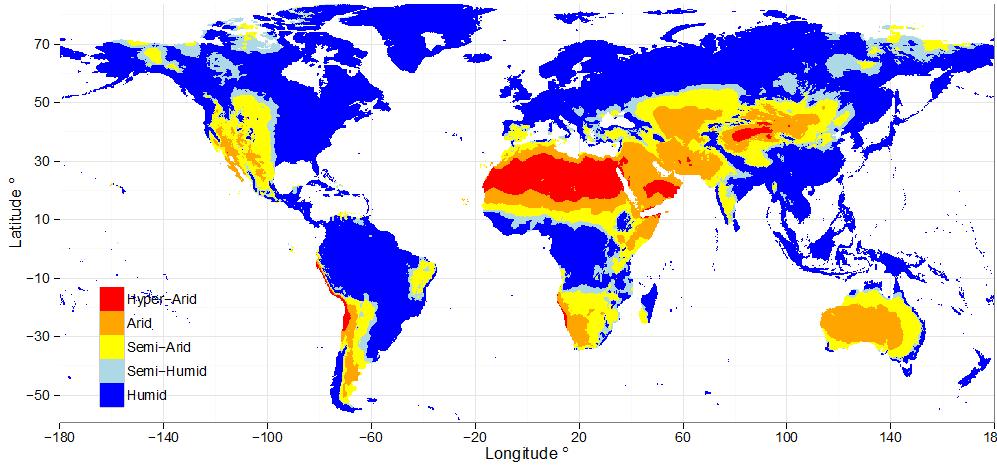
Figure 4 | Map of global distribution of climatic zones. (produced using data provided by CGIAR-CSI (link).
We found the skewed distribution of hydroclimatic sensitivity along the aridity gradient, i.e., rapid decline in sensitivity from the maximum sensitivity in semiarid region toward minimum sensitivity in arid regions, is very intriguing and important. This implies that shifts of the climate in semiarid regions to drier conditions will lead to rapid decay of sensitivity of productivity to hydroclimatic variations in these ecosystems.
Since 1970s, an overall drying trend in Southern Hemisphere semiarid regions has been noted (since 1950s for SE Australia; Cai and Cowan, 2013) coinciding with a poleward expansion of the subtropical dry zone that is partially attributable to anthropogenic climate change (Cai et al., 2012). This raises an important question of global biogeochemical significance: as to whether the large, terrestrial carbon sink noted in 2011, of which a significant fraction was apportioned to arid and semiarid regions of Australia, can be repeated with future hydroclimatic wet pulses if semiarid regions continue their shift to a drier climate?
This research was led by Prof. Alfredo Huete from C3’s Remote Sensing group and is jointly supported by Australian Research Council – Discovery Project (Huete) and NASA SMAP mission (Moran). Dr. Xuanlong Ma is a postdoctoral research fellow with expertise in remote sensing and terrestrial ecology. Please contact Dr. Xuanlong Ma (xuanlong.ma@uts.edu.au) and Prof. Alfredo Huete (alfredo.huete@uts.edu.au) for any further inquiry regarding this research.